
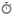
1. Reactive Species as Spatially and Temporally Controlled Signaling Entities
Reactive oxygen, nitrogen, and sulfur species (ROS/RNS/RSS) are increasingly understood as tightly regulated signaling mediators embedded within compartmentalized enzymatic networks rather than stochastic byproducts of oxidative metabolism. Hydrogen peroxide (H₂O₂), in particular, exhibits a unique physicochemical profile: moderate reactivity, relative stability compared to radical species, and selective oxidation of low pKa cysteine thiolates (Cys–S⁻), enabling signaling specificity under controlled microenvironmental conditions [1–3].
A defining principle of redox signaling is spatial restriction. H₂O₂ diffusion is not purely passive but is shaped by membrane permeability, aquaporin-mediated transport (peroxiporins), and rapid enzymatic consumption by peroxiredoxins (Prx), catalase, and glutathione peroxidases (GPx). This establishes steep intracellular redox gradients, particularly between mitochondria, endoplasmic reticulum, and cytosolic microdomains [4–6].
At the enzymatic source level, NADPH oxidase (NOX) complexes generate ROS in a regulated, receptor-coupled manner, while mitochondrial electron transport chain leakage (Complex I/III) contributes to basal redox tone. Importantly, these sources are not redundant but functionally specialized, producing discrete signaling pulses with distinct subcellular localization and temporal kinetics [7,8].
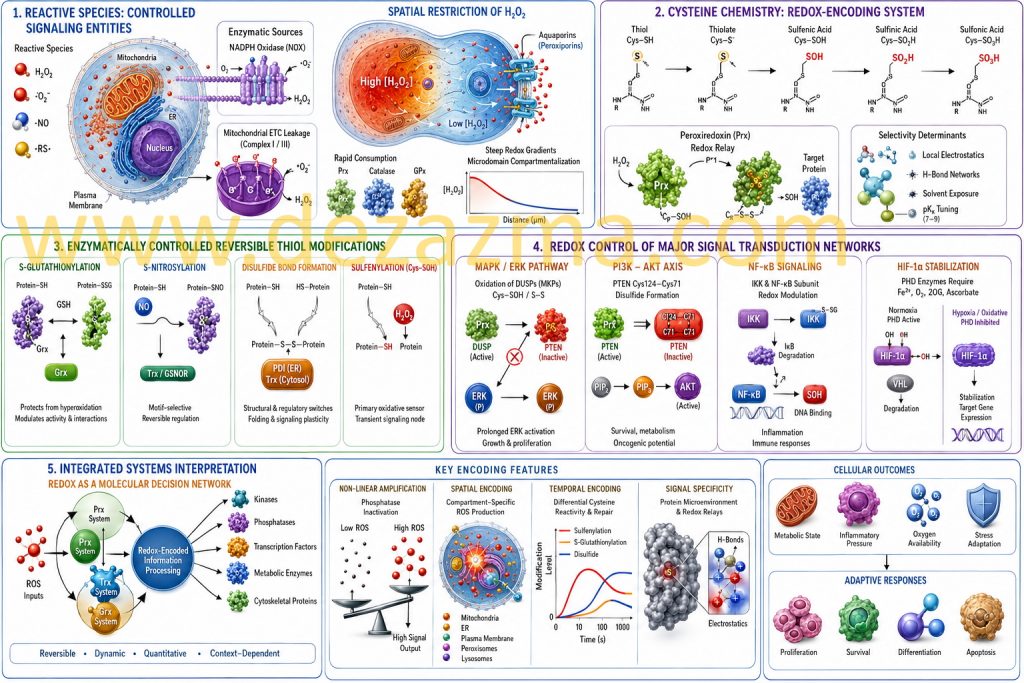
2. Cysteine Chemistry as a Redox-Encoding System
Protein cysteine residues operate as chemically programmable sensors due to their tunable pKa (typically 7–9), nucleophilicity, and susceptibility to stepwise oxidation states. The redox transition landscape includes:
Among these, sulfenylation (–SOH) is a critical kinetic intermediate, functioning as a transient signaling node rather than damage marker [9,10].
Thiol (Cys–SH) → thiolate (Cys–S⁻)
Sulfenic acid (Cys–SOH)
Sulfinic acid (Cys–SO₂H, partially reversible via sulfiredoxin in select proteins)
Sulfonic acid (Cys–SO₃H, generally irreversible physiological endpoint)
Selectivity in cysteine oxidation is not determined solely by solvent exposure but by local electrostatics, hydrogen bonding networks, and proximity to catalytic redox relays such as peroxiredoxins. In fact, peroxiredoxins function not only as antioxidants but also as H₂O₂ sensors and signal transducers via redox relay mechanisms, transferring oxidative equivalents to target proteins with high specificity [11,12].
3. Enzymatically Controlled Reversible Thiol Modifications
Redox-based post-translational modifications form an interconnected regulatory system:
S-glutathionylation arises through thiol–disulfide exchange or via sulfenic acid intermediates reacting with reduced glutathione (GSH). This modification protects cysteine residues from hyperoxidation while simultaneously altering protein conformation, enzymatic activity, and protein–protein interactions. It is dynamically regulated by glutaredoxin (Grx) systems [13].
S-nitrosylation is mediated through NO-derived nitrosating equivalents (e.g., N₂O₃, metal–NO complexes) and exhibits motif selectivity influenced by acid–base microenvironments and hydrophobic pockets. Denitrosylation is enzymatically regulated by thioredoxin and S-nitrosoglutathione reductase (GSNOR), enabling reversibility [14].
Disulfide bond formation acts as both structural and regulatory switches. In signaling contexts, dynamic disulfide exchange reactions are catalyzed by protein disulfide isomerases (PDI) in the ER and thioredoxin systems in the cytosol, ensuring controlled folding and signaling plasticity [15].
Sulfenylation (Cys–SOH) represents the primary oxidative sensing event. Its detection in living systems has revealed that many proteins undergo rapid, reversible sulfenylation bursts within seconds of ROS exposure, consistent with signal transduction rather than oxidative damage [16].
4. Redox Control of Major Signal Transduction Networks
MAPK/ERK Pathway
Redox regulation of MAPK signaling is primarily mediated through oxidation-sensitive phosphatases, particularly dual-specificity phosphatases (DUSPs/MKPs). Oxidation of catalytic cysteine residues within the active site (forming sulfenyl or disulfide states) inactivates phosphatase activity, thereby prolonging ERK phosphorylation and amplifying proliferative signaling outputs. This mechanism introduces a redox-dependent “gain control” layer over growth factor signaling [17].
PI3K–AKT Axis
The tumor suppressor PTEN contains a catalytically essential cysteine (Cys124) highly susceptible to reversible oxidation. Formation of a disulfide bond between Cys124 and Cys71 leads to conformational inactivation of PTEN phosphatase activity, resulting in increased PIP₃ accumulation and downstream AKT activation. This redox switch directly links oxidative tone to metabolic reprogramming, survival signaling, and oncogenic transformation potential [18].
NF-κB Signaling
NF-κB regulation exhibits multilayered redox sensitivity. At the upstream level, IKK complex activation is modulated by thiol oxidation and S-glutathionylation. At the transcriptional level, cysteine residues within the DNA-binding domain of NF-κB subunits (e.g., p50 Cys62) undergo redox modifications that alter DNA affinity. Thus, ROS functions both as an activator and modulator of transcriptional specificity depending on intensity and duration [19].
HIF-1α Stabilization Pathway
Hypoxia-inducible factor regulation is tightly coupled to redox state via prolyl hydroxylase domain (PHD) enzymes, which require Fe²⁺ and O₂ for activity. Oxidative modulation of iron center chemistry, as well as ascorbate and 2-oxoglutarate availability, suppresses hydroxylation of HIF-1α, preventing VHL-mediated degradation. This integrates oxygen sensing with redox signaling, allowing metabolic adaptation under hypoxic and oxidative stress conditions [20].
5. Integrated Systems Interpretation: Redox as a Molecular Decision Network
At a systems level, redox signaling constitutes a chemically encoded information-processing layer superimposed on classical phosphorylation networks.
The essential features include:
Non-linear amplification via reversible enzyme inactivation (phosphatases > kinases)
Spatial encoding via compartment-specific ROS production
Temporal encoding via differential cysteine reactivity and repair kinetics
Signal specificity via protein microenvironment and redox relays
Peroxiredoxin-based signal relays and thioredoxin/glutathione systems form a hierarchical buffering architecture that transforms oxidative inputs into graded, reversible signaling outputs. This enables cells to interpret redox fluctuations as quantitative parameters reflecting metabolic state, inflammatory pressure, and oxygen availability rather than binary stress signals [21,22].
References
Sies H. Hydrogen peroxide as a central redox signaling molecule. Redox Biol. 2017;11:613–619.
D’Autréaux B, Toledano MB. ROS signaling specificity. Nat Rev Mol Cell Biol. 2007;8:813–824.
Winterbourn CC. The biological chemistry of hydrogen peroxide. Methods Enzymol. 2013;528:3–25.
Rhee SG. Redox signaling: H₂O₂ as intracellular messenger. Exp Mol Med. 1999;31:53–59.
Miller EW et al. Aquaporin-mediated H₂O₂ transport. PNAS. 2010;107:15681–15686.
Veal EA et al. Peroxiredoxins as H₂O₂ sensors. J Biol Chem. 2007;282:26753–26759.
Bedard K, Krause KH. NOX family NADPH oxidases. Physiol Rev. 2007;87:245–313.
Murphy MP. Mitochondrial ROS signaling. Biochem J. 2009;417:1–13.
Poole LB, Nelson KJ. Cysteine oxidation mechanisms. Curr Opin Chem Biol. 2008;12:18–23.
Reddie KG, Carroll KS. Protein cysteine sulfenylation. Curr Opin Chem Biol. 2008;12:746–754.
Wood ZA et al. Peroxiredoxin catalytic cycle. Trends Biochem Sci. 2003;28:32–40.
Sobotta MC et al. Peroxiredoxin redox relays. PNAS. 2015;112:106–111.
Giustarini D et al. S-glutathionylation in signaling. Antioxid Redox Signal. 2004;6:345–360.
Foster MW et al. S-nitrosylation signaling. Trends Mol Med. 2009;15:391–404.
Hatahet F, Ruddock LW. Protein disulfide isomerase systems. FEBS J. 2009;276:194–201.
Paulsen CE, Carroll KS. Sulfenic acid biology. Chem Rev. 2013;113:4633–4679.
Kamata H et al. Redox regulation of MAPK signaling. Cell Signal. 1999;11:1–14.
Leslie NR. PTEN oxidation and PI3K signaling. Antioxid Redox Signal. 2007;9:1765–1774.
Morgan MJ, Liu ZG. ROS and NF-κB crosstalk. Cell Res. 2011;21:103–115.
Kaelin WG, Ratcliffe PJ. Oxygen sensing via HIF. Mol Cell. 2008;30:393–402.
Winterbourn CC. Redox biology complexity. Biochem J. 2013;461:1–10.
Jones DP. Redox sensing and signaling. Nat Rev Mol Cell Biol. 2006;7:392–401.

