استرس اکسیداتیو یکی از بنیادیترین مفاهیم در بیوشیمی مدرن و زیستشناسی محسوب میشود و به وضعیتی اطلاق میگردد که در آن تعادل دینامیک میان تولید گونههای فعال اکسیژن، نیتروژن و گوگرد (Reactive Oxygen, Nitrogen and Sulfur Species; ROS/RNS/RSS) و ظرفیت مکانیسمهای آنتیاکسیدانی سلول مختل شده و محیط سلولی به سمت شرایط اکسیدکننده سوق مییابد.
برخلاف دیدگاه سنتی که
ROS را صرفاً عوامل مخرب تلقی میکرد، امروزه مشخص
شده است که این مولکولها در غلظتهای فیزیولوژیک بهعنوان پیامرسانهای ثانویه
در تنظیم مسیرهای سیگنالینگ سلولی، تکثیر، تمایز، پاسخ ایمنی، تنظیم اپیژنتیک،
عملکرد میتوکندری و سازگاری متابولیک نقش اساسی ایفا میکنند. با این حال، افزایش
کنترلنشده تولید ROS یا
کاهش ظرفیت دفاع آنتیاکسیدانی منجر به آسیب تجمعی در سطح مولکولی، سلولی و بافتی
میشود.
مهمترین منابع درونسلولی تولید
ROS شامل کمپلکسهای I
و III زنجیره
انتقال الکترون میتوکندری، خانواده آنزیمهای
NADPH Oxidase (NOX)، زانتین اکسیداز، سیکلواکسیژنازها، لیپواکسیژنازها،
سیستم سیتوکروم P450،
پراکسیزومها و واکنشهای وابسته به یونهای فلزی واسطه مانند آهن و مس هستند.
در این میان، میتوکندری نهتنها منبع اصلی تولید سوپراکسید
محسوب میشود، بلکه بهعنوان هدف اولیه آسیب اکسیداتیو نیز شناخته میشود؛ بهطوریکه
اختلال در عملکرد میتوکندری میتواند یک چرخه معیوب از افزایش تولید ROS، کاهش تولید
ATP و تخریب ساختارهای سلولی را ایجاد کند.
در سطح مولکولی، استرس اکسیداتیو موجب پراکسیداسیون اسیدهای
چرب غیراشباع غشایی و تولید متابولیتهای واکنشپذیری نظیر مالوندیآلدئید (MDA)، 4-هیدروکسینوننال (4-HNE) و ایزوپروستانها میشود که
خود دارای فعالیتهای بیولوژیک و پاتوژنیک مستقل هستند. همچنین اکسیداسیون پروتئینها
منجر به تشکیل گروههای کربونیل، نیتروزیلاسیون تیروزین، اکسیداسیون باقیماندههای
سیستئین و متیونین و تغییرات ساختاری برگشتناپذیر میشود که عملکرد آنزیمها، گیرندهها
و پروتئینهای انتقالدهنده را مختل میکند. آسیب اکسیداتیو به DNA نیز شامل اکسیداسیون بازهای نوکلئوتیدی،
تشکیل 8-هیدروکسی-2′-دئوکسیگوانوزین (8-OHdG)،
شکست رشتههای DNA و
افزایش ناپایداری ژنومی است که در بروز سرطان و فرآیند پیری نقش محوری دارند.
در پاسخ به این چالش، سلولها شبکهای پیچیده از مکانیسمهای دفاعی را توسعه دادهاند که شامل آنتیاکسیدانهای آنزیمی نظیر سوپراکسید دیسموتاز (SOD)، کاتالاز (CAT)، گلوتاتیون پراکسیداز (GPx)، پراکسیردوکسینها (Prx) و تیوردوکسینها (Trx) و نیز آنتیاکسیدانهای غیرآنزیمی همچون گلوتاتیون احیاشده (GSH)، کوآنزیم Q10، ویتامین C، ویتامین E، اسید اوریک، بیلیروبین و پلیفنولهای گیاهی است. تنظیم بیان بسیاری از این سیستمهای دفاعی توسط فاکتور رونویسی Nrf2 انجام میشود که امروزه یکی از مهمترین اهداف درمانی در پزشکی مولکولی محسوب میگردد.
از منظر بالینی، شواهد گسترده نشان میدهد که استرس اکسیداتیو
در پاتوژنز طیف وسیعی از بیماریها از جمله دیابت ملیتوس، سندرم متابولیک،
آترواسکلروز، بیماریهای قلبی–عروقی، سرطان، بیماری آلزایمر، پارکینسون، بیماریهای
خودایمنی، نارسایی کلیوی، بیماریهای کبدی و اختلالات مرتبط با سالمندی نقش اساسی
دارد. علاوه بر این، تعامل پیچیده میان استرس اکسیداتیو، التهاب مزمن، اختلال
عملکرد میتوکندری و تغییرات اپیژنتیک بهعنوان یک محور مرکزی در زیستشناسی بیماریهای
مزمن شناخته میشود.
بیوشیمی میکروبیوم را میتوان بهعنوان مطالعهی شبکههای پیچیدهی تبادل متابولیکی و شیمیایی میان میکروارگانیسمهای همزیست و میزبان تعریف کرد؛ شبکهای که در آن جریان مداوم متابولیتها، آنزیمها و سیگنالهای مولکولی، فنوتیپ فیزیولوژیک میزبان را بهصورت مستقیم و غیرمستقیم تنظیم میکند. در این چارچوب، میکروبیوم نه صرفاً یک مجموعهی منفعل از میکروبها، بلکه یک «اندام متابولیک افزوده» در نظر گرفته میشود که ظرفیتهای بیوشیمیایی میزبان را گسترش میدهد.
در سطح متابولیکی، میکروبیوتای روده نقش محوری در تخمیر پلیساکاریدهای غیرقابل هضم (dietary fibers) دارد که منجر به تولید اسیدهای چرب کوتاهزنجیر شامل استات، پروپیونات و بوتیرات میشود. این ترکیبات از طریق اتصال به گیرندههای G-protein coupled receptors نظیر GPR41 (FFAR3) و GPR43 (FFAR2) در سلولهای اپیتلیال، سلولهای ایمنی و بافت چربی، مسیرهای تنظیم انرژی، التهاب و هموستاز گلوکز را تعدیل میکنند. بوتیرات علاوه بر نقش انرژیزایی برای کولونوسیتها، بهعنوان یک مهارکنندهی هیستونداستیلاز (HDAC inhibitor) نیز عمل کرده و از طریق تنظیم اپیژنتیکی، بیان ژنهای مرتبط با التهاب و تکامل سلولهای T تنظیمی را تغییر میدهد.
در محور متابولیسم اسیدهای صفراوی، میکروبیوتا با دِکونژوگاسیون و تبدیل اسیدهای صفراوی اولیه به فرمهای ثانویه، ترکیب پویای مخزن صفراوی را تغییر میدهد. این متابولیتها از طریق گیرندههایی مانند FXR (Farnesoid X Receptor) و TGR5 بر متابولیسم لیپیدها، حساسیت به انسولین و پاسخهای التهابی اثر میگذارند. بنابراین، میکروبیوم بهطور غیرمستقیم در تنظیم محور کبد–روده و هموستاز کلسترول نقش دارد.
از منظر مسیر تریپتوفان، میکروبیوتا با تبدیل این اسیدآمینه به ایندولها و مشتقات آن، سیگنالهایی تولید میکند که گیرندهی AhR (Aryl hydrocarbon Receptor) را فعال میسازند. این مسیر در تنظیم سد اپیتلیال روده، تولید پپتیدهای ضد میکروبی و پاسخهای ایمنی ذاتی اهمیت کلیدی دارد. اختلال در این مسیر با بیماریهای التهابی روده و دیسبیوزیس مرتبط است.
در سطح متابولیسم نیتروژن و کولین، تبدیل کولین و کارنیتین غذایی به TMA توسط میکروبیوتا و سپس اکسیداسیون کبدی آن به TMAO (Trimethylamine N-oxide)، نمونهای از تعامل میکروبیوم–میزبان در پاتوژنز بیماریهای قلبی–عروقی محسوب میشود. این مسیر نشان میدهد که متابولیتهای میکروبی میتوانند اثرات سیستمیک دور از محل تولید خود ایجاد کنند.
از نظر روششناسی، تحلیل بیوشیمی میکروبیوم بر تلفیق چندین فناوری پیشرفته استوار است:
Metagenomics برای شناسایی ظرفیت ژنتیکی جامعه میکروبی
Metatranscriptomics برای بررسی فعالیت ژنی در شرایط واقعی
Metabolomics برای پروفایلسازی متابولیتهای کوچک
Fluxomics و مدلسازی متابولیکی در مقیاس ژنومی (GEMs) برای بازسازی جریانهای بیوشیمیایی در شبکههای متابولیک
ترکیب این دادهها امکان ایجاد مدلهای سیستمزیستی (systems biology) از محورهای عملکردی مانند روده–مغز، روده–کبد و روده–ایمنی را فراهم میسازد.
در سطح بالینی، این دانش پایهی مفاهیمی مانند پزشکی دقیق مبتنی بر میکروبیوم (microbiome-based precision medicine)، طراحی نسل جدید پروبیوتیکها و پستبیوتیکها، و نیز توسعهی رویکردهای فارماکومتابولومیک در پیشبینی پاسخ دارویی را تشکیل میدهد. همچنین نقش آن در بیماریهای متابولیک (دیابت نوع ۲، چاقی)، اختلالات نورودژنراتیو، بیماریهای التهابی و سرطانها بهطور فزایندهای مورد توجه قرار گرفته است.
این سایت برای آموزش بیوشیمی بالینی و تبادل آرا بین علاقمندان این علم طراحی شده است. نام سایت شامل دو کلمه است. دز مخفف دزفول بعنوان یکی از قدیمی ترین مراکز علم و تمدن در ایران و آز که مخفف آزمایشگاه است و اشاره به نقش تحقیقات در توسعه علم بیوشیمی دارد.

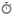


{kind=link}