
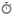
محقق : زینب حمزه لو-علوم آزمایشگاهی
 DNA مارپیچی دو رشته ای
DNA مارپیچی دو رشته ای
از نیمه ی قرن 20 تا کنون که DNA به عنوان حامل اطلاعات ژنتیکی شناخته شد تلاش برای مشخص شدن اساس ساختاری ذخیره ی اطلاعات تشدید شد . ولی چند تصور اشتباه پیشرفت را کند کرد . جدی ترین آنها مشاهدات آزمایشگاهی اشتباهی بود که DNA را دارای مقادیر یکسانی از هر یک از چهار نوکلئوتید (دی اکسی آدنوزین، دی اکسی گوانوزین، دی اکسی سیتیدین و دی اکسی تیمیدین ) می دانستند .
آزمایشات صناعی و پراش اشعه ی X منجر به پذیرفتن ساختار خطی پلی نوکلئوتید برای DNA شد . با این حال ساختار سه بعدی DNAهنوز به صورت یک معما باقی ماند . سه بخش از اطلاعات برای استنتاج ساختار DNA لازم بود . اولی تعیین می کرد که DNA دارای مقادیر یکسان از چهار نوکلئوتید نبوده و در موجودات مختلف دارای مقادیر متغییری است با این حال مشخص شد که مقدار دی اکسی آدنوزین همیشه برابر با دی اکسی تیمیدین است و مقدار دی اکسی گوانوزین همیشه برابر با دی اکسی سیتیدین می باشد . (dA با dT و dG با dC ) .
دومین مسئله که ناشی از نتایج اشعه ی X بود نشان داد که DNA دارای ساختار مارپیچ دو رشته ای است و تقارن موجود بیان می کرد که رشته های پلی نوکلئوتیدی به صورت غیر هم سو با یکدیگر جهت دهی شده اند . مطلب سوم این بود که نوکلئوبازها به حالتهای توتومری کتو و آمین بودند نه به حالتهای انول و ایمین . توتومرها ، ایزومرهای مولکولی هستند که فقط موقعیت یک اتم هیدروژن در آنها متفاوت است . هر چهار نوکلئوباز دارای دو یا تعداد بیشتری از حالتهای توتومری هستند که با یکدیگر در حال تعادلند . ارجاع نادرست ساختار توتومری غالب موجب شده بود که محققان سعی کنند جفت بازهایی را با استفاده از الگوهای اتصالات هیدروژنی نادرست ایجاد کنند .
ساختاری که در سال 1953 توسط واتسون و کریک برای DNA مارپیچ دورشته ای پیشنهاد شد به دلیل سادگی و تقارن آنها مورد توجه قرار گرفت . علاوه بر این ، این ساختار پیشنهادی همه ی اطلاعات ساختاری موجود در DNA را توصیف کرده و منجر به ارائه ی فرضیه هایی درباره ی مکانیسم نگهداری اطلاعات ژنتیکی گردید . مارپیچ دو رشته ای واتسون- کریک می تواند به صورت دو رشته ی پلی نوکلئوتیدی مارپیچی راست گرد حول یک محور مشترک در نظر گرفته شود . رشته ها از طریق اتصال پیوندهای هیدروژنی که در لبه های هیدروفیلیک بازها تشکیل می شوند فشرده می شوند . گروه های H-N نوکلئوبازها دهنده های پیوند هیدروژنی خوبی بوده و جفت الکترونهای روی اکسیژن های دارای هیبرید SP2 گروه های C=O و نیتروژنهای گروه های – N = گیرنده های پیوند هیدروژنی هستند . جفت شدن زمانی صورت می گیرد که یک گیرنده و یک دهنده در موقعیتی قرار گرفته باشند که پیوند هیدروژنی تشکیل دهند این پیوندها بین واحدهای پورین در یک رشته و واحدهای پیریمیدین در رشته دیگر صورت گرفته و جفت شدن دهنده ها و گیرنده های پیوند هیدروژنی دونوع جفت باز تشکیل می دهد : آدنین – تیمین و گوانین – سیتوزین .
یک پیامد مستقیم از خصوصیات پیوندهای هیدروژنی این است که DNA دو رشته ای ( dsDNA ) باید دارای نسبت هایی از نوکلئوزیدها باشد که متناسب با مشاهدات آزمایشگاهی باشد ( dT=dA وdC=dG ) . ارتباط بین بازهای رشته های مخالف در مارپیچ دورشته ای به عنوان مکمل بودن توصیف می شود . بازها مکمل یکدیگرند چون هر نوکلئوباز یک رشته از طریق پیوندهای هیدروژنی با باز رشته ی دیگر جفت می شود . برای مثال هر آدنینی که به طرف محور مارپیچ دورشته ای بیرون زده است یک تیمین باید در رشته ی مخالف قرار گرفته و با ایجاد پیوند هیدروژنی به صورت کامل فضای بین رشته ها را پر نماید . نه سیتوزین و نه گوانین به صورت کامل و دقیق ، نمی توانند فضای موجود در عرض آدنین را که از طریق پیوند هیدروژنی موجود در عرض رشته ها پر می شود تکمیل کنند .
این خصوصیات پیوندهای هیدروژنی است که توالی بازیک رشته کاملا با رشته ی دیگر مکمل است . بخش خارجی مارپیچ دورشته ای از اسکلت فسفات – قند اجزای آن تشکیل شده است . پلی نوکلئوتیدها به دلیل قطبیت ذاتی ساختارهای غیر متقارن هستند . دو رشته در جهت مخالف ردیف می شوند . اگر دو نوکلئوباز مجاور در یک رشته – مثلاً تیمین و سیتوزین – در جهت 3َ <- 5َ به هم متصل شوند ، نوکلئوبازهای مکمل آنها در رشته ی دیگر یعنی آدنین و گوانین در جهت 5َ <- 3َ به هم متصل می شوند . پیچش های بین دو رشته ی نا هم سو ، ساختاری ایجاد می کند که دارای دو شکاف بین اسکلت قند – فسفات است . شکاف بزرگ خیلی پهن تر از شکاف کوچک است . این تفاوت حاصل ساختار هندسی جفت بازهاست : پیوندهای گلیکوزیدی بین بازها و پنتوز اسکلت مستقیماً در جهت مخالف هم قرار نگرفته اند بلکه به سمت شکاف کوچک جا به جا شده اند . توالی نوکلئوتیدی DNA می تواند بدون جدا کردن مارپیچ دو رشته ای ، از طریق این شکاف ها تشخیص داده شود .
هر باز همیشه اتم های یکسانی را داخل هر شکاف مارپیچ دو رشته ای نشان می دهد . بنابراین این اتم ها یک ابزار مهم برای تشخیص توالی خاص DNA از طریق پروتئین و مولکولهای به وجود می آورند .

