
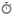
کاتابولیسم اسکلت کربنی اسیدهای آمینه
اسکلت کربنی اسیدهای آمینه میتواند به کربوهیدراتها و چربیها تبدیل گردد. به اسیدهای آمینهای که به استیل کوآ و استواستیل کوآ تبدیل شوند، کتوژنیک گفته میشود. در مقابل به اسیدهای آمینهای که به پیروات، – کتوگلوتارات، سوکسینیل کوآ، فومارات و اگزالواستات تبدیل میشوند، گلوکوژنیک اتلاق میگردد. دو اسید آمینه لوسین و لیزین فقط کتوژنیک هستند. چهار اسید آمینه ایزولوسین، فنیل آلانین، تریپتوفان و تیروزین توانایی تبدیل شدن به گلوکز و استیل کوآ یا استواستیل کوآ را دارند که به آنها کتوژنیک و گلوکوژنیک میگویند.
اسیدهای آمینه از راههای مختلف میتوانند وارد مسیرهای سنتز گلوکز یا چربیها شوند. حذف شدن نیتروژن عامل آمین معمولاً اولین واکنش کاتابولیک است. در اثر این ترانس آمیناسیون ترکیبات واسطی ایجاد میشوند که میتوانند مسیرهای متابولیسمی مختلفی را طی کنند.
آسپاراژین و اسید آسپارتیک: آسپاراژین و اسید آسپارتیک در اثر ترانس آمیناسیون تولید اگزالواستات میکنند. اگزالواستات از مواد واسط در چرخه کربس و مسیر گلوکونئوژنز است، به همین سبب این دو اسید آمینه میتوانند در پایان به گلوکز تبدیل شوند.
گلوتامین و اسید گلوتامیک: مسیر کاتابولیسم گلوتامین و اسید گلوتامیک تا حد زیادی مشابه مسیر آسپاراژین و اسید آسپارتیک میباشد با این تفاوت که در این مسیر به جای اگزالواستات، آلفا- کتوگلوتارات تولید میشود. ماده اخیر از مواد اخیر از مواد واسط در چرخه کربس بوده و به همین سبب این دو اسید آمینه توانایی تبدیل شدن به گلوکز را دارند.
پرولین: پرولین جزء اسیدهای آمینهای است که مستقیماً ترانس آمینه نمیگردد. این اسید آمینه ابتدا به دهیدروپرولین تبدیل میشود که پس از چند واکنش آنزیمی به گلوتامات و آلفا- کتوگلوتارات تبدیل میشود. به همین دلیل پرولین در خانواده اسیدهای آمینه گلوکوژنیک طبقهبندی میگردد.
آرژینین: آرژینین تحت اثر آنزیم آرژیناز ابتدا به اسید آمینه اورنیتین تبدیل میگردد. اورنیتین در اثر ترانس آمیناسیون به گلوتامات- گاما- سمی آلدئید تبدیل میشود. ادامه مسیر مشابه روندی است که در کاتابولیسم پرولین رخ میدهد.
هیستیدین: در اثر دآمیناسیون هیستیدین مادهای به نام اوروکانات تشکیل میگردد. این ماده پس از طی چند واکنش آنزیمی ابتدا به گلوتامات سپس به آلفا- کتوگلوتارات تبدیل میشود. در مسیر کاتابولیسم هیستیدین مادهای به نامN – فورمیمینوگلوتامات (figlu) تولید میشود. در صورتی که بدن با کمبود اسیدفولیک مواجه شود، میزان دفع این ماده از طریق ادرار افزایش مییابد.
سرین: سرین عمدتاً توسط آنزیم سرین هیدروکسی متیل ترانسفراز به گلیسین تجزیه میگردد.
گلیسین نیز به طور عمده در یک مسیر غیرقابل برگشت توسط مجموعه گلیسین سنتتاز شکسته میگردد.
همچنین سرین توسط آنزیم سرین دهیدراتاز میتواند به پیروات تبدیل شود.
سیستئین: سیستئین از دو مسیر کاتابولیزه میشود. محصول نهایی در هر دو مسیر پیروات است.
الف) مسیر اکسیداتیو مستقیم:
ب) مسیر ترانس آمیناسیون
سیستین در پستانداران اغلب توسط آنزیم سیستئین ردوکتاز به سیستئین تبدیل شده و سیستئین نیز طی یکی از روندهای فوق کاتابولیزه میگردد.
ترئونین: ترئونین توسط آنزیم آدولاز به استالدئید و گلیسین شکسته میشود. گلیسین میتواند از طریق سرین در پایان به پیروات تبدیل شود. همچنین استالدئید در انتها به استیل کوآنزیم A تبدیل میگردد. به همین دلیل ترئونین جزء اسیدهای آمینه گلوکوژنیک طبقهبندی میشود، زیرا توانایی تبدیل شدن به پیروات و استیل کوآنزیم A را دارد.
تیروزین: در بین اسیدهای آمینه کاتابولیسم تیروزین یکی از پیچیدهترین روندهای کاتابولیک است. در طول این مسیر اختلالات آنزیمی زیادی به چشم میخورد. تیروزینمی و آلکاپتونوری از شایعترین این اختلالات هستند.
فنیل آلانین: در شرایط طبیعی مهمترین مسیر متابولیسمی فنیل آلانین از طریق تبدیل شدن به تیروزین است. در بیماری فنیل کتونوری (PKU) به علت نقص آنزیم فنیل آلانین هیدروکسیلاز این مسیر مسدود شده و راههای جانبی فعالتر میگردند.
لیزین: لیزین جزء اسیدهای آمینهای است که ازتهای آن در ترانس آمیناسیون شرکت نمیکنند. لیزین پس از طی چندین واکنش آنزیمی در انتها به گلوتاریل کوآنزیم A تبدیل میشود که این ماده نیز پس از چند واکنش به و تجزیه میشود.
تریپتوفان: مسیر کاتابولیسم تریپتوفان از برخی جهات شبیه مسیر کاتابولیسم لیزین میباشد. اولین آنزیم طی روند کاتابولیسم تریپتوفان آنزیم تریپتوفان اکسیژناز یا تریپتوفان پیرولاز است. نقص این آنزیم سبب بیماری هارت ناپ میگردد. محصول نهایی کاتابولیسم تریپتوفان نیز و است.
متیونین و اسیدهای آمینه شاخهدار: سوکسینیل کوآنزیم A محصول نهایی مهم کاتابولیسم متیونین، ایزولوسین و والین میباشد. سوکسینیل کوآنزیم A از مواد واسط در چرخه کربس است. به همین دلیل این اسیدهای آمینه توانایی تبدیل شدن به گلوکز را دارند. از بین اسیدهای آمینه شاخهدار لوسین در انتها به بتا ـ هیدروکسی بتا ـ متیل گلوتاریل کوآنزیم A (HMG-CoA) تبدیل میشود. HMG-CoA ماده واسط در سنتز کلسترول و اجسام کتونی است. به همین علت لوسین اسید آمینهای است که فقط کتوژنیک میباشد.
اختلال در کاتابولیسم اسیدهای آمینه شاخهدار سبب بیماری شربتافرا (Maple Syrup) میگردد. بوی ادرار این افراد شبیه بوی شیره درخت افرا میشود.
متیونین همچنین میتواند به کمک ATP، S– آدنوزیل متیونین (SAM) را تولید کند که مهمترین دهنده ریشه متیل در بدن است.
هوموسیستئین: این اسید آمینه در ساختمان پروتئینها به کار نرفته است و از متابولیسم متیونین حاصل میگردد. در طبقهبندی اسیدهای آمینه جزء اسیدهای آمینه گوگرددار تقسیمبندی میشود.
در مسیر متابولیسم متیونین ـ هوموسیستئین علاوه بر آنزیمهای فوق به آنزیم متیل تتراهیدروفولات ردوکتاز نیز نیاز میباشد. ویتامین ، اسیدفولیک و در این مسیر نقش کوآنزیمی دارند.
افزایش هوموسیستئین میتواند علت ارثی یا اکتسابی داشته باشد. نقص آنزیمی به ویژه آنزیمهای سیستاتیونین سنتتاز و متیل تتراهیدروفولات ردوکتاز از علل ارثی افزایش هوموسیستئین خون است. کمبود ویتامینهای ، و اسیدفولیک از عوامل اکتسابی افزایش هوموسیستئین خون میباشند.
نیتریک اکسید (NO): NO ملکولی با خواص منحصر به فرد است. این مولکول به راحتی از غشاء عبور میکند و اعمال گوناگونی را در بدن انجام میدهد. NO در بدن از اسید آمینه L– آرژینین حاصل میگردد. یکی از مهمترین نقشهای نیتریک اکسید عمل انبساط عروق (وازودیلاتوری) است.
دکربوکسیلاسیون اسیدهای آمینه
از دکربوکسیلاسیون اسیدهای آمینه، آمینهایی با اعمال تخصص یافته حاصل میگردند. کوآنزیم تمام آنزیمهای دکربوکسیلاسین اسیدهای آمینه فسفات پیریدوکسال (شکل فعال ویتامین ) است. در اینجا تعدادی از مهمترین محصولات حاصل از دکربوکسیلاسیون اسیدهای آمینه را مرور میکنیم.
دکربوکسیلاسیون اسید آسپارتیک: در میکروارگانیسمها بتا ـ آلانین از آلفا ـ دکربوکسیلاسیون اسیدآسپارتیک حاصل میگردد.
مسیرهای تجربه اسیدهای آمینه
مسیرهای کاتابولیسم اسیدهای آمینه، در مجموع، به طور طبیعی تنها 10% تا 15% تولید انرژی را در بدن انسان شامل میگردند؛ این مسیرها به اندازه گلیکولیز و اکسیداسیون اسیدهای چرب فعال نمیباشند. جریان این مسیرها نیز برحسب تعادل بین نیاز به اسیدهای آمینه در فرآیندهای بیوسنتتیک و موجود بودن یک اسید آمینه خاص، به میزان قابل توجهی متفاوت میباشد. 20 مسیر کاتابولیک تنها به پنج محصول ختم میشوند که تمامی آنها وارد چرخه اسید سیتریک میگردند. از اینجا به بعد، اسکلتهای کربنی ممکن است در گلوکونئوژنز یا کتوژنز شرکت نموده و یا به طور کامل به یا اکسیده گردند.
تمامی یا قسمتی از اسکلت کربنی ده اسید آمینه نهایتاً به استیلکوآ تجزیه میشوند. پنج اسید آمینه به – کتوگلوتارات، چهار اسید آمینه به سوکسینیل کوآ، دو اسید آمینه به فومارات و دو اسید آمینه به اگزالواستات تبدیل میگردند. مسیرهای خاص هر 20 اسید آمینه در اشکالی خلاصه شدهاند و هر کدام از آنها در یک نقطه ورودی اختصاصی وارد چرخه اسید سیتریک میشوند. در این دیاگرامها، اتمهای کربن ورودی به چرخه اسید سیتریک، نشان داده شدهاند. توجه نمایید که تعدادی از اسیدهای آمینه بیش از یک بار نمایش داده شدهاند که سرنوشتهای مختلف قسمتهای متفاوت اسکلت کربنی آنها را منعکس مینماید. به جای بررسی هر مرحله از هر مسیر کاتابولیسم اسیدهای آمینه، بعضی از واکنشهای آنزیمی که از نظر مکانیسم و اهمیت پزشکی ذکر آنها ارزش دارد، به طور خاص مورد برسی قرار میگیرند.
بسیاری از کوفکتورهای آنزیمی، نقشهای مهمی را در کاتابولیسم اسیدهای آمینه ایفاء مینمایند.
انواع مختلفی از نوآراییهای شیمیایی جالب توجه در مسیرهای کاتابولیک اسیدهای آمینه رخ میدهد. شروع بحث با مسیرهایی مفید خواهد بود که کلاسهایی از واکنشها را نشان میدهند که تکرار شده و نقش کوفکتورهای آنزیمی خود را معرفی مینمایند. تاکنون یک کلاس مهم، یعنی واکنشهای ترانسآمیناسیون نیازمند پیریدوکسال فسفات، را مورد توجه قرار دادهایم. نوع معمول دیگری از واکنشها در کاتابولیم اسیدهای آمینه، شامل انتقالات یک کربنه میباشد که به کمک یکی از کوفکتورهای بیوتین، تتراهیدروفولات و S– آدنوزیل متیونین به انجام میرسند. این کوفکتورها، گروههای یک کربنه را با وضعیتهای مختلف اکسیداسیون منتقل مینمایند: بیوتین کربن را در اکسیدهترین حالت، یعنی ، منتقل مینماید. تتراهیدروفولات گروههای یک کربنه را در حالات اکسیداسیون حد واسط و گاهی به صورت گروههای متیل انتقال میدهد؛ و S– آدنوزیل متیونین گروههای متیل، احیاء شدهترین وضعیت کربن، را انتقال میدهد. دو کوفکتور اخیر اهمیت خاصی در متابولیسم اسیدهای آمینه و نوکلئوتیدها دارند.
تتراهیدروفولات () از پترین استخلاف شده (5- متیل پترین)، پارا ـ آمینوبنزوات و قسمتهای گلوتامینی تشکیل شده است. این کوفکتور در باکتریها سنتز میگردد. شکل اکسیده این ترتیب، یعنی فولات، برای پستانداران به عنوان ویتامین مطرح بوده که طی دو مرحله توسط دیهیدروفولات ردوکتاز به تتراهیدروفولات تبدیل میگردد. گروه یک کربنه انتقالی، در هر کدام از سه حالات اکسیداسیون، به ازت 5 یا ازت 10 و یا هر دو، متصل میگردد. احیاء شدهترین شکل کوفکتور، یک گروه متیل را حمل نموده، شکل اکسیدهتر حامل ی گروه متیلن بوده و اکسیدهترین حالت یک گروه متنیل، فرمیل و یا فورمیمینو را منتقل مینماید. اکثر اشکال تتراهیدروفولات قابل تبدیل به یکدیگر بوده و به عنوان دهندههای واحدهای یک کربنه در انواع مختلفی از واکنشهای متابولیکی شرکت مینمایند. منبع اصلی واحدهای یک کربنه برای تتراهیدروفولات، کربنی است که در هنگام تبدیل سرین به گلیسین برداشت شده و تولید ، متیلن تتراهیدروفولات مینمایند.
هر چند تتراهیدروفولات میتواند یک گروه متیل را بر روی ازت 5 خود حمل نماید، این پتانسیل انتقال گروه متیل برای اکثر واکنشهای بیوسنتتیک کافی نمیباشد. S– آدنوزیل متیونین (adoMet)، کوفکتور ترجیحی برای انتقالات بیولوژیکی گروه متیل میباشد. این کوفکتور از ATP و متیونین در حضور متیونین آدنوزیل ترانسفراز سنتز میگردد. این واکنش از این نظر غیرمعمول است که اتم سولفور نوکلئوفیل متیونین به کربن َ5 قسمت ریبوزی ATP، بجای یکی از اتمهای فسفر، حمله مینماید. در نتیجه، تریفسفات رها شده که بر روی آنزیم به Pi و PPi شکسته شده و خود PPi بعداً توسط پیروفسفاتاز معدنی تجزیه میگردد؛ از اینرو، طی این واکنش سه پیوند، شامل دو پیوند پرانرژی، شکسته میشود. تنها واکنش شناخته شده دیگر که طی آن تریفسفات از ATP جدا میگردد، در سنتز ویتامین رخ میدهد.
انتقال گروه متیل از S– آدنوزیل متیونین به یک گیرنده، همراه با تولید S– آدنوزیل هموسیستئین میباشد (مرحله2) که بعداً به هموسیستئین و آدنوزین تجزیه میگردد (مرحله 3). با انتقال یک گروه متیل به هموسیستئین در واکنشی با حضور متیونین سنتاز، متیونین مجدداً تولید میگردد (مرحله 4) و برای تکمیل یک چرخه متیل فعال شده، متیونین مجدداً به S– آدنوزیل متیونین تبدیل میشود.
یک شکل معمول متیونین سنتاز در باکتریها از متیل تتراهیدروفولات به عنوان دهنده متیل استفاده مینماید. شکل دیگر این آنزیم که در باکتریها و پستانداران وجود دارد، از متیل تتراهیدروفولات و یا متیل کوبالامین مشتق از ویتامین بهره میبرد. این واکنش و نوآرایی L– متیل مالونیل کوآ به سوکسینیل کوآ، تنها واکنشهای شناخته شده وابسته به کوآنزیم در پستانداران میباشند. در کمبود ویتامین ، بعضی از علائم را میتوان نه تنها با تجویز ، بلکه همچنین با فولات، تخفیف داد. گروه متیل کوبالامین از متیل تتراهیدروفولات مشتق میگردد. از آنجایی که واکنش تبدیل شکل ، متیلن به شکل متیل تتراهیدروفولات غیرقابل برگشت میباشد، در صورت وجود کمبود کوآنزیم برای سنتز متیل کوبالامین، گیرنده دیگری برای گروه متیل متیل تتراهیدروفولات وجود نخواهد داشت و فولاتهای متابولیک به شکل بدام میافتند. پنهان شدن فولات به این شکل، ممکن است علت بعضی از علائم بیماری کمخونی کشنده ناشی از کمبود ویتامین باشد. هر چند نمیدانیم که آیا این تنها اثر کمبود ویتامین است.
تتراهیدروبیوپترین، کوفکتور دیگر متابولیسم اسیدهای آمینه، مشابه قسمت پترین تتراهیدروفولات بوده ولی در انتقالات یک کربنه شرکت نداشته و در واکنشهای اکسیداسیون همکاری دارد. این نحوه عمل را در بحث تجزیه فنیلآلانین مورد توجه قرار خواهیم داد.
ده اسید آمینه به استیلکوآ تجزیه میگردند.
اسکلت کربنی ده اسید آمینه میتوانند بعد از تولید استیلکوآ وارد چرخه اسید سیتریک شده و یا در سنتز اسیدهای چرب مورد استفاده قرار گیرند. پنج اسید آمینه از این ده مورد، از طریق پیروات به استیلکوآ تبدیل میگردند که میتوانند در سنتز گلوکز نیز شرکت کنند. پنج اسید آمینه دیگر مستقیماً به استیلکوآ و یا به استواستیل کوآ تجزیه شده که بعداً به استیلکوآ تبدیل میشوند.
پنج اسید آمینهای که به پیروات تبدیل میشوند، شامل آلانین، تریپتوفان، سیستئین، سرین و گلیسین میباشند. در بعضی از موجودات، ترئونین نیز به استیلکوآ تجزیه میشود؛ همانطور که بعداً شرح داده خواهد شد، ترئونین در انسان به سوکسینیل کوآ تجزیه میگردد. آلانین بدنبال ترانس آمیناسیون با – کتوگلوتارات، مستقیماً تولید پیروات نموده و شکسته شدن زنجیر جانبی تریپتوفان همراه با تولید آلانین و بنابراین پیروات میباشد. تبدیل سیستئین به پیروات طی دو مرحله، شامل برداشت اتم سولفور و ترانس آمیناسیون، صورت میپذیرد. سرین دهیدراتاز تبدیل سرین به پیروات را کاتالیز مینماید. هر دو گروه – هیدروکسیل و – آمینو سرین در این واکنش وابسته به پیریدوکسال فسفات، برداشت میگردد.
گلیسین دارای دو مسیر میباشد. در مسیر اصلی باکتریایی، گلیسین با افزودن یک گروه هیدروکسی متیل توسط آنزیم به سرین تبدیل میگردد. این واکنش توسط سرین هیدروکسی متیل ترانسفراز کاتالیز شده و نیاز به کوآنزیمهای تتراهیدروفولات و پیریدوکسال فسفات دارد. در مسیر دوم که در حیوانات غالب است، گلیسین متحمل تجزیه اکسیداتیو به ، و یک گروه متیلن () میگردد. این واکنش که بسادگی قابل برگشت میباشد، توسط گلیسین سنتاز کاتالیز شده و نیاز به تتراهیدروفولات به عنوان گیرنده گروه متیلن دارد. در این مسیر تجزیه اکسیداتیو، دو اتم کربن گلیسین وارد چرخه اسید سیتریک نمیشوند. یک کربن به صورت از دست رفته و اتم کربن دیگر به گروه متیلن ، متیلن تتراهیدروفولات، یک دهنده گروه یک کربنه در بعضی مسیرهای بیوسنتتیک خاص، تبدیل میشود.
قسمتهایی از اسکلت کربنی شش اسید آمینه، شامل تریپتوفان، لیزین، فنیل آلانین، تیروزین، لوسین و ایزولوسین، تولید استیلکوآ یا استواستیل کوآ و یا هر دو را مینمایند؛ سپس ترکیب اخیر به استیلکوآ تبدیل میگردد. بعضی از مراحل نهایی موجود در مسیرهای تجزیهای لوسین، لیزین و تریپتوفان مشابه اکسیداسیون اسیدهای چرب میباشد. مسیرهای تجزیهای دو اسید آمینه از این شش مورد، نیاز به توجه خاص دارند.
تجزیه تریپتوفان، پیچیدهترین مسیر در کاتابولیسم اسیدهای آمینه در بافتهای حیوانی میباشد؛ قسمتهایی از تریپتوفان (شش کربن از کل کربنهای آن)، طی دو مسیر متفاوت، یکی از طریق پیروات و دیگری از طریق استواستیلکوآ، تولید استیلکوآ مینمایند. بعضی از ترکیبات واسط موجود در کاتابولیسم این اسیدهای آمینه، پیشسازهایی برای سنتز بیومولکولهای دیگر، شامل نیکوتینات به عنوان پیشساز NAD و NADP در حیوانات، سروتونین به عنوان یک نوروترانسمیتر در مهرهداران و اندولاستات به عنوان یک فاکتور رشد در گیاهان، میباشند. جزئیات بعضی از این مسیرهای بیوسنتتیک در فصل 22 بیشتر شرح داده میشود.
بررسی تجزیه فنیلآلانین اهمیت دارد، زیرا همانطور که در قسمت پائین شرح داده میشود، نقصهای ژنتیکی آنزیمهای شرکت کننده در این مسیر منجر به بیماریهای متعدد ارثی در انسان میشوند. فنیلآلانین و تیروزین به عنوان محصول اکسیداسیون آن (هر دو 9 کربنه)، به دو قطعه تجزیه میشوند که هر دو میتوانند وارد چرخه اسید سیتریک گردند: چهار اتم کربن به استواستات تبدیل شده که به استواستیل کوآ و بنابراین استیل کوا تبدیل میگردد و قطعه چهار کربنه دیگری به صورت فومارات آزاد میشود. بنابراین 8 کربن از 9 کربن این دو اسید آمینه وارد چرخه اسید سیتریک شده و کربن باقیمانده به صورت از دست میرود. فنیل آلانین، بعد از هیدروکسیلاسیون به تیروزین، به عنوان پیش ساز نورترانسمیتر دوپامین و هورمونهای نوراپی نفرین و اپی نفرین مترشحه از قسمت مرکزی غده فوق کلیوی، نیز عمل مینماید. ملانین، رنگدانه سیاه پوست و مو، نیز مشتقی از تیروزین میباشد.
آمینواسیدهای شاخهدار، استیل COA، استواستات یا پروفیونیل COA تولید میکنند.
تجزیه آمینواسیدهای شاخهدار بوسیله واکنشهایی انجام میشود که قبلاً در چرخه اسید سیتریک و اکسایش اسیدهای چرب با آنها آشنا شدیم. لوسین ترانس آمینه شده و به کتواسید معادل خود یعنی کتوایزو کاپروئات تبدیل میگردد. این کتواسید کمپلکس آنزیمی کتواسید دهیدروژناز مؤثر بر کتواسیدهای شاخهدار به طور اکسایشی به ایزو والریل COA دکربوکسیله میشود.
کتواسیدهای والین و ایزولوسین، دو آمینواسید دیگر با زنجیره آلیفاتیک شاخهدار، و نیز کتوبوتیرات مشتق شده از متیونین از سوبستراهای دیگر این آنزیم هستند. دکربوکسیله شدن اکسایشی این کتواسیدها، مشابه تبدیل پیرووات به استیل COA و کتوگلوتارات به سوکسینیل COA است. کتواسید دهیدروژناز مؤثر بر کتواسیدهای شاخهدار یک کمپلکس چند آنزیمی، همولوگ با پیرووات دهیدروژناز و کتوگلوتارات دهیدروژناز به شمار میآید. در حقیقت، اجزاء این آنزیمها که شکل اکسایش یافته لیپوآمید را باز تولید میکنند، یکسان هستند.
ایزووالریل COA مشتق شده از لوسین، دهیدروژنه شده تا متیل کروتونیل COA تولید گردد. این عمل اکسایش، به وسیله آنزیم ایزووالریل COA دهیدروژناز کاتالیز میشود. پذیرنده هیدروژن نظیر واکنش مشابه در اکسایش اسیدهای چرب که بوسیله استیل COA دهیدروژناز کاتالیز میگردد، FAD است. سپس متیل گلوتاکونیل COA از طریق کربوکسیله شدن متیل کروتونیل COA و یا هزینه هیدرولیزیک مولکول ATP تشکیل میگردد. همانطوریکه انتظار میرود، مکانیسم کربوکسیله کردن توسط متیل کروتونیل COA کربوکسیلاز شبیه مکانیسم پیرووات کربوکسیلاز و استیل COA کربوکسیلاز میباشد.
سپس متیل گلوتاکونیل COA جهت تشکیل 3- هیدروکسی 3- متیل گلوتاریل COA هیدراته میگردد که این متابولیت به استیل COA و استواستات سکسته میشود. این واکنش قبلاً در ارتباط با تشکیل اجسام کتونی از اسیدهای چرب مورد بحث قرار گرفته است.
مسیرهای تجزیهای والین و ایزولوسین شبیه مسیر تجزیه لوسین میباشد. بعد از ترانس آمینه شدن و دکربوکسیله شدن اکسایشی جهت تشکیل مشتقات CoA، واکنشهای بعدی شبیه اکسایش اسیدهای چرب هستند. ایزولوسین، استیل CoA و پروپیونیل CoA ایجاد میکند در حالیکه والین و پروپیونیل CoA تولید میکند. تجزیه لوسین، والین و ایزولوسین، تأیید نکتهای است که قبلاً مورد تأکید قرار داده بودیم. تعداد واکنشهای موجود در متابولیسم زیاد است، اما انواع واکنشها نسبتاً کم است. تجزیه لوسین، والین و ایزولوسین توصیف جالب توجهی از سادگی و ظرافت اصول متابولیسم را فراهم میآورد.

